- Introduzione e diffusione in Europa
- Morfologia
- Ecologia
- Impatti
- Tossicità
- Tossicità per gli animali
- Tossicità per l’uomo
- Bibliografia tematica
- Lavori generali e informazioni botaniche varie (morfologia, ecologia, cariologia…)
- Arrivo in Europa, diffusione, areale distributivo
- Comportamento invasivo, strategie di invasività, Risk Assessment
- Impatti, intossicazione animali e uomo
- Tossicità in miele e polline
- Aspetti tossicologici: alcaloidi, inquinamento alimentare, normative
- Sitografia
- Bibliografia (ordine alfabetico)
Introduzione e diffusione in Europa
Il senecione o senecio sudafricano (Senecio inaequidens DC.) è una specie erbacea perenne, appartenente alla famiglia delle Asteraceae. Originaria del Sudafrica è giunta in Europa accidentalmente verso la metà del XX secolo, probabilmente sotto forma di seme in partite di lana grezza.
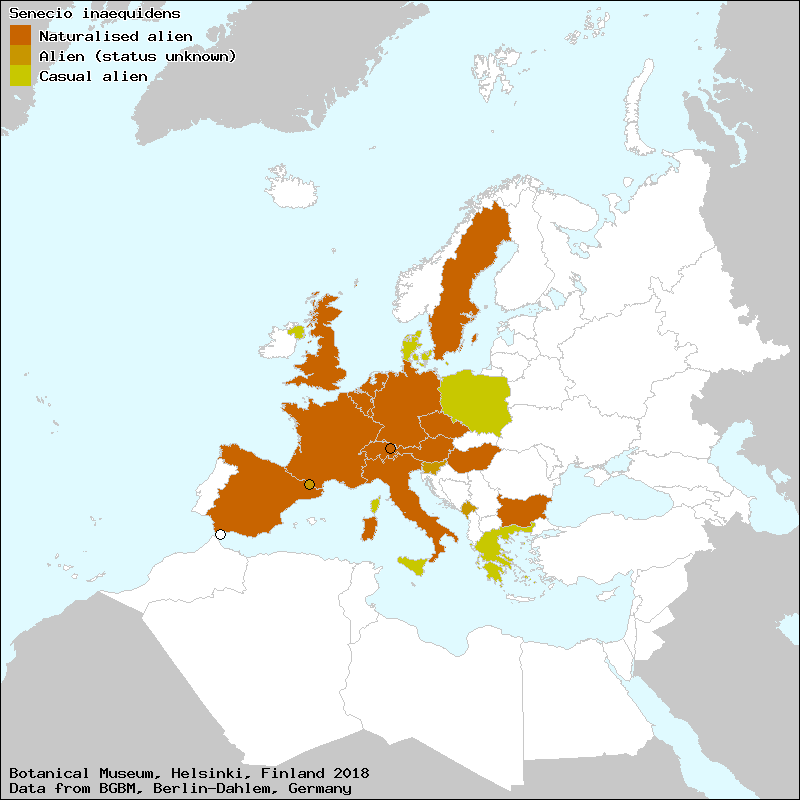
Il suo arrivo e la successiva rapida diffusione nell’Europa centrale sono ben documentati: dopo il primo ritrovamento, risalente al 1889 e riguardante l’area di Hannover in Germania, la pianta è stata segnalata nella prima metà del ‘900 in Belgio, Gran Bretagna, Francia, Olanda e Italia, solitamente in località connesse con processi industriali di lavorazione della lana. In tempi più recenti la sua capacità di diffusione è andata aumentando fino a renderla presente in buona parte dei Paesi europei.
Euro+Med – gennaio 2021
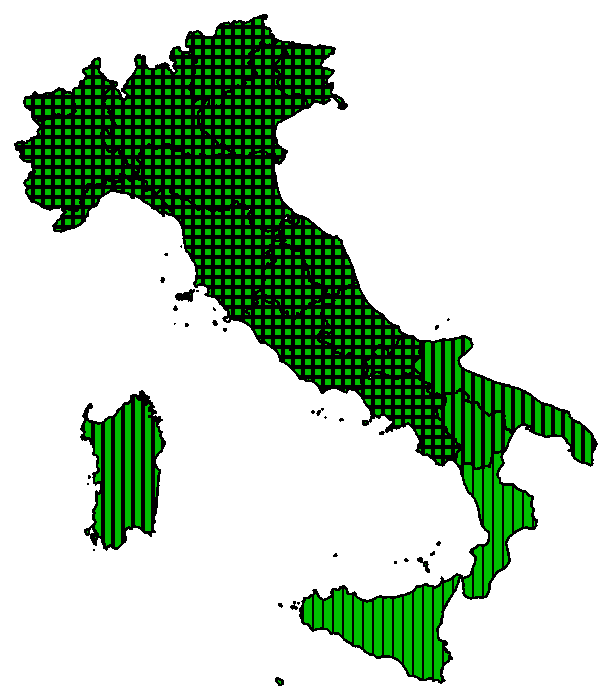
In Italia è stato ritrovato per la prima volta nel 1947, nel veronese, dove tutt’ora risulta abbondante (Anzalone 1976). Nel 1982 Pignatti (1982) lo indicava per le sole regioni centro-settentrionali della penisola, ma già nel 2005 era noto in tutte le regioni italiane con l’esclusione di Puglia e Calabria (Conti et al. 2005). Oggi risulta presente su tutto il territorio nazionale (Portale della Flora d’Italia 2021).
Da Portale Flora d’Italia – gennaio 2021
Morfologia
Il senecio sudafricano può vivere tra i 5 e 10 anni. Ogni singola pianta produce numerosi fusti, legnosi e prostrati alla base, eretti, ramificati e di consistenza erbacea più in alto, che possono raggiungere e spesso superare i 60 cm d’altezza (non è raro incontrare individui di un metro). Le foglie, sessili e abbraccianti, sono acuminate, strette e allungate (2-4 mm di larghezza e 6-7 cm di lunghezza), con bordo irregolarmente dentato (da cui l’epiteto specifico “inaequidens”). I capolini sono terminali, di diametro compreso tra 1,5 e 2,5 cm e di colore giallo. In base a quanto riportato in letteratura, ogni pianta può arrivare a portare circa 80-100 capolini, tuttavia nella nostra esperienza abbiamo riscontrato anche oltre 600 capolini su una singola pianta. La fioritura del senecio è molto vistosa ed estremamente prolungata, protraendosi da aprile fino a dicembre-gennaio.



Un singolo individuo produce in media circa 10.000 frutti (acheni) ma può arrivare a produrne fino a 30.000. Ogni achenio misura circa 2 mm, è estremamente leggero e dotato di un pappo piumoso che ne facilita la dispersione anemofila. È in grado di produrre semi già dal primo anno di vita. Riguardo alla vitalità dei semi non c’è molta chiarezza in letteratura: contrariamente alle opinioni espresse su alcuni report ufficiali (NOBANIS 2006; EPPO 2004) in cui si ipotizza una persistenza della seed bank nel suolo per 30-40 anni, indagini più recenti evidenziano che il 25% dei semi sepolti rimangono vitali dopo un anno, ma i semi sulla superficie del terreno perdono vitalità dopo 6 mesi (López-García & Maillet 2005; USDA 2005; EPPO 2006; Curtaz et al. 2011). Riesce a moltiplicarsi anche per via vegetativa attraverso l’emissione di radici dai fusti che toccano terra.
L’impollinazione è entomofila e i fiori sono visitati da numerosi insetti generalisti tra cui principalmente ditteri, lepidotteri e imenotteri.




Ecologia
Si tratta di una specie estremamente rustica e adattabile, capace di occupare e diffondersi rapidamente in numerosi ambienti anche molto diversi. L’ampia tolleranza ecologica e la capacità di crescita estremamente rapida gli consentono di insediarsi nelle aree di pianura come alle quote più elevate: in Val d’Aosta sono state rilevate piante oltre i 1700 m di quota, in Lombardia (Bresciano) fino a 1500 m mentre nel suo areale di origine si spinge fino a 2850 m. Colonizza sia zone umide sia ambienti secchi, tanto su suoli acidi quanto su quelli basici (pur preferendo tendenzialmente i substrati calcarei). Generalmente predilige gli ambienti disturbati insediandosi di preferenza in aree antropizzate come campi, pascoli, vigneti, ambienti ruderali (macerie, aree abbandonate, base dei muri ed edifici). Le vie di comunicazione come strade, sentieri, ferrovie e scarpate stradali rappresentano senza dubbio un ambiente d’elezione per il senecio i cui semi riescono a sfruttare i fenomeni di turbolenza dovuti al passaggio degli automezzi. Spesso si ritrova in aree più naturali come incolti sassosi, rupi e greti dei fiumi. Talvolta anche in prati più o meno radi o in giovani arbusteti, mai in boschi più o meno densi e ombrosi.
La produzione di un elevatissimo numero di semi rappresenta un chiaro punto di forza che contribuisce a rendere questa specie più competitiva di altre nel colonizzare nuovi ambienti. Il principale vettore per la sua diffusione è il vento: i semi, provvisti di pappo, possono essere facilmente trasportati anche su lunghe distanze (EPPO 2006). Monty et al. (2008) stabiliscono in 100 m la distanza entro cui viene depositato il 99.8% dei semi, con un picco massimo intorno ai 5,2 m. Tuttavia, questa sperimentazione si basa su velocità del vento relativamente modesta (5 m/sec) e bassa turbolenza e gli stessi autori ammettono che in condizioni ambientali diverse (soprattutto con forti raffiche di vento), le distanze possono essere sensibilmente diverse. Anche gli animali, soprattutto con pelliccia, possono fungere da vettori nel trasferimento dei semi. La sua diffusione inoltre è favorita dal passaggio di automezzi e treni che dislocano i semi lungo le direttrici principali. Infine, può spostarsi accidentalmente attraverso la movimentazione di terra, materiali da costruzione o macchinari agricoli.
Recenti indagini hanno dimostrato che le popolazioni presenti nell’area nativa sono sia diploidi che tetraploidi, mentre quelle presenti in Europa appartengono solamente al citotipo tetraploide (Monty et al. 2010). La resistenza al freddo varia molto a seconda del citotipo: la sopravvivenza all’inverno è praticamente nulla per gli individui diploidi, bassa per i tetraploidi nativi ed è più elevata nei tetraploidi presenti nei territori invasi. Secondo vari Autori sembra quindi che in Europa si sia avuta una leggera divergenza rispetto alle popolazioni del Sud Africa che ha portato ad un maggiore adattamento verso la resistenza al secco e al freddo, fino alla diversificazione, in alcune aree, di due fasi fenologiche con due periodi di fioritura (tarda primavera e inverno) e quindi con la possibilità di due produzioni di semi per anno (USDA 2005).
Impatti
L’azione di disturbo di Senecio inaequidens nei confronti di singole specie non è ancora ben documentata e, nonostante in letteratura si trovino opinioni contrastanti, è evidente che una sottrazione di spazio nei confronti di entità endemiche o con areale contenuto può rappresentare un serio problema. L’elevata competitività e l’ingente tasso riproduttivo ne fanno invece una evidente minaccia per diverse comunità vegetali. Dal punto di vista ecologico è indiscutibile che la vistosa e prolungata fioritura (protratta anche per 8-9 mesi all’anno che comprende anche i mesi invernali) costituisca una rilevante alterazione del paesaggio naturale. Inoltre va osservato che recenti studi evidenziano in Senecio inaequidens uno degli invasori in Europa con più rapida capacità di colonizzazione di nuovi ambienti.
La sua elevata plasticità fenotipica suggerisce un notevole potenziale bio-ecologico non ancora sfruttato, che potrebbe determinare la migrazione verso nuove zone climatiche e il possibile adattamento a situazioni al momento ritenute sfavorevoli (Monty & Mahy, 2009; Monty et al. 2013); quindi una specie che potrebbe avvantaggiarsi dai potenziali scenari di cambiamento climatico.
Sotto l’aspetto economico il senecio africano apporta danni diretti come specie infestante dei coltivi e dei pascoli contribuendo a ridurne il valore commerciale. Reinhardt et al. (2003), pur ammettendo la difficoltà di quantificarne il valore monetario, evidenziano anche danni indiretti dovuti alla necessità di sostenere costi per la manutenzione stradale.
Attualmente, l’aspetto di maggior rilievo riguarda comunque l’impatto sanitario.
Tossicità
La pericolosità del senecio sudafricano è principalmente legata alla presenza di alcaloidi pirrolizidinici (Pyrrolizidine Alkaloids, abbreviati in PAs), fitocomposti naturali che si differenziano in oltre 350 molecole – di cui circa la metà tossiche – e che possono provocare fenomeni di avvelenamento del bestiame e dell’uomo. Si stima che circa 6.000 specie di piante nel mondo possano contenere tali alcaloidi, che si concentrano principalmente in alcune famiglie di angiosperme: Boraginaceae, Asteraceae e Fabaceae (genere Crotalaria). In natura i PAs svolgono un ruolo importante in relazione ai meccanismi di difesa di piante e insetti. I fitofagi evitano le piante ad elevato tenore in PAs. Alcuni insetti, invece, se ne cibano e attraverso questi composti tossici si difendono dai loro antagonisti. Altri insetti, come le falene ad esempio, trasformano i PAs in feromoni che hanno un ruolo fondamentale nell’accoppiamento (Kast et al. 2010).
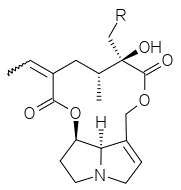
(R=H per Senecionina, R=OH per Retrorsina)
Nelle Asteraceae gli alcaloidi sono principalmente sintetizzati nelle radici, distribuiti lungo le fibre in tutta la pianta. Dalla pianta possono facilmente passare ad animali e uomo attraverso varie modalità. Il contenuto di PAs negli alimenti e nei mangimi dipende da numerosi fattori, compresi specie e organo della pianta, raccolta, conservazione e procedure di estrazione.
Retrorsina e Senecionina sono due dei principali PAs contenuti in Senecio inaequidens. Sono tra gli alcaloidi pirrolizidinici maggiormente tossici esistenti in natura e sono sufficienti tra i 30 e i 50 milligrammi per ogni kg di peso corporeo per accusare sintomi gravissimi o addirittura mortali (Tabella 1).
| ALCALOIDI PIRROLIZIDINICI | LD50 (mg/kg) | RELATIVE PIANTE PRODUTTRICI |
| Retrorsina | 36 | Senecio cineraria, S. erucifolius, S. inaequidens, S. jacobaea, S. vulgaris |
| Senecionina | 50 | Petasites hybridus, Senecio cineraria, S. erraticus, S. erucifolius, S. inaequidens, S. jacobaea, S. nemorensis, S. petasitis, S. squalidus, S. subalpinus, S. viscosus, S. vulgaris, Tussilago farfara |
| Eliotrina | 296 | Heliotropium curassavicum, H. europaeum, H. supinum |
| Licopsamina | >1000 | Anchusa officinalis, Borago officinalis, Symphytum asperum, S. officinale |
Il problema della presenza negli alimenti e nei mangimi, degli alcaloidi pirrolizidinici ha sollevato di recente una forte preoccupazione anche a livello Comunitario, tanto che l’Autorità Europea per la Sicurezza Alimentare (EFSA) ha pubblicato un parere scientifico su questa tematica. Gli esperti scientifici del Gruppo sui Contaminanti nella Catena Alimentare (CONTAM) hanno individuato la presenza negli alimenti e nei mangimi di un certo numero di PAs, importanti in quanto potenziali contaminanti e hanno concluso che esiste una possibile preoccupazione sanitaria per alcuni forti consumatori di miele, il solo alimento per il quale fossero disponibili dati sui livelli di PAs. Inoltre hanno confermato che determinati alcaloidi pirrolizidinici possono agire sull’uomo da cancerogeni genotossici (EFSA 2011). In letteratura sono evidenziati numerosi casi di intossicazione, sia animale che umana, in diverse parti del mondo. Testimonianze di malatie epatiche causate sul bestiame dall’ingestione di erbe contenenti alcaloidi pirrolizidinici (Senecio jacobaea e genere Crotalaria) esistono già dai primi del ‘900 (Wiedenfield 2011b). Tra i casi più recenti, che hanno dato impulso a nuove indagini, è da menzionare invece la morte di numerose vacche nel 2004 in Sudafrica, intossicate a seguito dell’ingestione di Senecio inaequidens in un pascolo pesantemente invaso (Dimande et al., 2007). Anche le intossicazioni umane da alcaloidi sono descritte già nei primi decenni del ‘900: risale al 1920 l’intossicazione in Sudafrica causata dal consumo di pane con semi contaminati da specie del genere Senecio. I casi più gravi di intossicazione si sono rilevati in Afghanistan e Tagikistan: nel 1975-1976, 8000 persone sono rimaste intossicate dal consumo di cereali contaminati da Heliotropium popovii subsp. gillianum e, similmente, 4000 persone sono stati ricoverate nel 1992 a causa di grano inquinato dalla presenza di Heliotropium lasiocarpum (10-12). Sempre H. lasiocarpum è responsabile di un episodio di intossicazione avvenuto alla fine del 1940 nell’ex Unione Sovietica, sempre a carico di pane contaminato (Wiedenfield 2011b).
Gli attuali metodi per analizzare il contenuto di PAs in alimenti e mangimi si basano essenzialmente su gascromatografia-spettrometria di massa (GC-MS) e su cromatografia liquida-spettrometria di massa tandem (LC-MS/MS). In Italia solo poche ditte altamente specializzate sono in grado di effettuare analisi per il rilevamento dei PAs all’interno degli alimenti.
Tossicità per gli animali

Il bestiame può ingerire il senecio al pascolo o con il fieno fornito nella razione. Gli animali al pascolo tendono a scartare la pianta, a causa del suo gusto amaro, che però scompare con l’affienamento, aumentando il rischio di ingestione del senecio nel fieno. Poiché l’essiccazione non elimina la tossicità del senecio, ingerendo la pianta l’animale accumula la tossina nel proprio organismo. Gli alcaloidi pirrolizidinici vengono assorbiti nel tratto gastrointestinale e agiscono principalmente nel fegato, dove liberano molecole tossiche che si accumulano e danneggiano progressivamente l’organo interessando, a volte, anche il cuore e i polmoni. Non esiste, attualmente, un trattamento per contrastare l’avvelenamento da PAs. I sintomi da avvelenamento sono difficilmente identificabili, anche perché, trattandosi generalmente di tossicità cronica, può manifestarsi diversi mesi dopo l’ingestione. Il bestiame avvelenato può presentare sintomi quali inappetenza, perdita di peso, diarrea, problemi neurologici, letargia ecc. Il livello di tossicità del senecio sudafricano per il bestiame può variare in relazione alla specie, all’età, al sesso e allo stato fisiologico e nutrizionale degli animali. La bibliografia sull’argomento analizza principalmente il problema dell’avvelenamento dei cavalli, più sensibili all’intossicazione da Senecio inaequidens, anche perché spesso pascolano in aree marginali, dove il senecio trova condizioni ottimali per la propria diffusione. In realtà, anche le parcelle destinate alla produzione di fieno, se invase da Senecio inaequidens, possono rappresentare una via di avvelenamento non trascurabile. La dose letale nei cavalli si raggiunge con un’ingestione di circa 300 g al giorno di pianta (espressa come sostanza secca) per un periodo di 50 giorni. Questo valore corrisponde al 3-5% del peso vivo dell’animale. Oltre ai cavalli è segnalata un’alta sensibilità anche per bovini, suini e galline. In generale, invece, ovicaprini, tacchini e ungulati selvatici (cervi) sembrano essere più tolleranti. Le capre e le pecore al pascolo a inizio primavera si nutrono delle rosette fogliari e raramente manifestano problemi. In diversi studi, la resistenza delle pecore all’intossicazione da alcaloidi è attribuita all’attività batterica del rumine, che consentirebbe la degradazione della molecola. Gli animali giovani sono più sensibili degli individui adulti. L’ingestione di elevate quantità di senecio in un breve tempo provoca un’intossicazione più rapida che l’assunzione di quantità inferiori per un tempo più lungo.
Tossicità per l’uomo
L’uomo può essere intossicato dagli alcaloidi del senecio sudafricano per imperizia nella raccolta di specie selvatiche per la produzione di rimedi naturali o per consumo alimentare di piante selvatiche. Tralasciando queste vie dirette d’intossicazione, l’uomo può entrare in contatto con gli alcaloidi pirrolizidinici, mediante il consumo di latte, miele, uova, carne e integratori alimentari di origine vegetale, prodotti da animali intossicati. Trattandosi spesso di specie vegetali gradite alle api, tracce di PAs possono ritrovarsi anche all’interno del miele. Gli effetti sull’uomo riguardano principalmente il fegato, con occlusione delle vene epatiche, emorragie necrotiche, sviluppo di fibrosi o cirrosi, a seconda che si tratti di tossicità acuta, sub-acuta o cronica. La sensibilità è maggiore nei soggetti più giovani e negli individui di sesso maschile.
Tra gli alimenti che possono contenere PAs in tracce, il miele è stato senza dubbio uno di quelli più studiati negli ultimi anni (Gallina, 2014).
Attualmente nell’Unione Europea non vigono valori limite per i PAs negli alimenti. Numerosi Paesi, tuttavia, stanno discutendo sulla opportunità di fissare una concentrazione massima consentita per la commercializzazione dei prodotti alimentari.
In alcuni casi si applicano già limiti massimi per l’assunzione di farmaci vegetali che contengono PAs. In Germania dal 1992 esiste un’ordinanza federale che regolamenta la vendita di prodotti medicinali a base di piante produttrici di AP con le seguenti dosi ammesse (Gallina, 2014):
- 1 µg al giorno per un periodo massimo di assunzione orale di 6 settimane;
- 100 µg al giorno per uso esterno e per un periodo massimo di 6 settimane;
- 0.1 µg al giorno per più di 6 settimane consecutive di assunzione orale;
- 10 µg al giorno per uso esterno, per un massimo di 6 settimane consecutive;
Il Ministero della Sanità Pubblica tedesco raccomanda di non superare la dose di 0.007 μg PAs/kg peso corporeo al giorno (correspondente a 0.42 μg PAs al giorno per una persona di 60 kg).
In Austria è ammessa la vendita di poche piante o preparati a base di piante produttrici di PAs. Tuttavia tali prodotti possono essere commercializzati solo previa analisi “condotta con le più recenti metodiche per l’analisi di PAs disponibili in letteratura”, e attestante la totale assenza di PAs nel prodotto.
In Olanda il contenuto totale di PAs e di PAs N-OX prodotti medicinali non può superare 1µg/kg o 1µg/l; per non incorrere in effetti cancerogeni ha stabilito un ADI (Acceptable Daily Intake) di 0,1µg/kg di peso corporeo al giorno (Gallina 2014).
Secondo Swissmedic i PAs tossici presenti nei farmaci vegetali devono essere dichiarati e il loro dosaggio deve essere tale da non superare l’assunzione giornaliera di 0,1 µg.
Se le direttive relative ai farmaci vegetali si applicassero agli alimenti, al miele ad esempio, supponendo che questo prodotto venga consumato in porzioni da 20 g al giorno, risulterebbe una concentrazione massima consentita compresa tra 5 e 50 µg di PAs per chilo di miele (Kastl et al. 2010).
A titolo di esempio, nel miele uniflorale di Echium vulgare (erba viperina comune) sono state riscontrate concentrazioni di PAs fino a 3.900 µg/kg (Kastl et al. 2010). Contenuto del tutto simile di PAs è stato riscontrato nel miele di Senecio jacobea (Edgar et al. 2002).
Uno studio tedesco ha analizzato oltre 200 campioni di miele distribuito nei supermercati di tutto il mondo. Nel 9% di essi è stato possibile riscontrare concentrazioni di PA comprese tra 19 e 120 µg /kg. Molti dei campioni positivi contenevano polline di Echium vulgare. Dübecke et al. (2011) hanno rilevato PAs in 696 campioni di miele da supermercato. Nel 94 per cento dei casi il tenore in PAs andava da 1 µg/kg a 267 µg/kg. Nell’88 per cento dei campioni analizzati i tenori in PAs erano inferiori a 50 µg/kg. Soltanto il 12% per cento dei campioni conteneva più di 50 µg/kg di PA.
Un recente lavoro di Martinello et al. (2014) ha preso in esame 70 campioni di miele in commercio nei supermercati italiani e provenienti da varie parti del mondo. Nel 64% dei casi sono stati riscontrati PAs, con concentrazioni più elevate per Lycopsamine, Intermedine, Echimidine, e Senecionine (Figura 9). La concentrazione media di PAs è risultata la seguente: 1,35 µg/kg nei mieli italiani, 3,14 µg/kg in quelli provenienti dall’UE e 17,45 µg/kg in quelli misti tra quelli europei e extracomunitari. I campioni più contaminati contenevano una concentrazione variabile tra 40 e 172 µg/kg. Il più contaminato ha raggiunto il valore di 243 µg/kg (Martinello et al. 2014).
Bibliografia tematica
Lavori generali e informazioni botaniche varie (morfologia, ecologia, cariologia…)
- Bornkamm R., 2002 – On the phytosociological affiliations of an invasive species Senecio inaequidens in Berlin. Preslia, 74(4): 395-407.
- Bouvet D., Selvaggi A., Siniscalco C., Soldano A., 2013 – Senecio inaequidens DC. In: Bouvet D. (ed.), Piante esotiche invasive in Piemonte. Riconoscimento, distribuzione, impatti. Museo Regionale di Scienze Naturali, Torino, 352 pp.
- Chichiricco G., Frizzi G., Tammaro R., 1979 – Numeri cromosomici per la Flora Italiana. Informatore Botanico Italiano, 11: 3-35.
- Curtaz A., Talichet M., Barni E., Bassignana M., Masante D., Pauthenet Y., Siniscalco C., 2011 – Specie esotiche invasive e dannose nei prati di montagna. Caratteristiche, diffusione e metodi di lotta. Institut Agricole Régional, Aosta.
- EPPO, 2006 – EPPO data sheet on Invasive Plants. Senecio inaequidens. Web version 2006-02-01 – doc 05-11836.
- Harland S C, (1954) The genus Senecio as a subject for cytogenetic investigation. Proceedings of the Botanical Society of the British Isles, 1, 256.
- Heger, T., Böhmer, H.J., 2006 – NOBANIS – Invasive Alien Species Fact Sheet – Senecio inaequidens. – From: Online Database of the European Network on Invasive Alien Species – NOBANIS www.nobanis.org. Date of access 20/08/2014.
- Lafuma L., Balkwill K., Imbert E., Verlaque R., Maurice,S., 2003 – Ploidy level and origin of the European invasive weed Senecio inaequidens (Asteraceae). Plant Systematics & Evolution. 243(1-2): 59-72.
- USDA, 2005 – Weed Risk Assessment for S. inaequidens DC. and S. madagascariensis Poir (Asteraceae). United States Department of Agriculture, Animal and Plant Health Inspection Service.
- Vanparys V., Cawoy V., Mahaux O., Jacquemart A.-L., 2011 – Comparative study of the reproductive ecology of two co-occurring related plant species: the invasive Senecio inaequidens and the native Jacobaea vulgaris. Plant Ecology and Evolution 144 (1): 3–11.
- Vanparys V., Meerts P., Jacquemart A.-L., 2008 – Plant–pollinator interactions: comparison between an invasive and a native congeneric species. Acta Oecologica, 34: 361-369.
- Wittenberg R. (ed.), 2005 – An inventory of alien species and their threat to biodiversity and economy in Switzerland. CABI Bioscience Switzerland Centre report to the Swiss Agency for Environment, Forests and Landscape.
Arrivo in Europa, diffusione, areale distributivo
- Anzalone B., 1976 – Il Senecio inaequidens DC. in Italia. Giornale Botanico Italiano, 110: 437-438.
- Carrara Pantano A., Tosco U., 1959 – Una nuova avventizia per la flora italiana: Senecio reclinatus L. f. di origine sud-africana, nella campagna veronese. Mem. Mus. Civ. St. Nat. Verona, Verona, 7: 151-157.
- Ernst W.H.O., 1998 – Invasion, dispersal and ecology of the South African neophyte Senecio inaequidens in the Netherlands: from wool alien to railway and road alien. Acta Botanica Neerlandica, 47(1), 131-151.
- Heger T., Böhmer H.J., 2005 – The invasion of Central Europe by Senecio inaequidens DC. – A complex biogeographical problem. Erdkunde, 59, 34-49.
- Hilliard O.M, Burtt B.L., 1975 – Notes on some plants of Southern Africa chiefly from Natal: IV. Notes from the Royal Botanic Garden Edinburgh, 34: 73-100.
- Michez J.M., 1994 – Senecio inaequidens. From the wool to the vine. Phytoma, 468, 39-41.
- Muller S., Maillet J., 2004 – Senencio inaequidens DC. Le séneçon du Cap. In: Muller S. (coord.), Plantes invasives en France. Muséum national d’Histoire naturelle, Paris, 168 pp.
- Often A., 1997 – Senecio inaequidens DC. and Solidago rugosa Mill. found as ruderals in Oslo. Blyttia, 55(3): 141-144.
- Sans F.X.; Garcia-Serrano H.; Afán I., 2004 – Life-history traits of alien and native senecio species in the Mediterranean region. Acta Oecologica, 2 (3): 167-178.
- Sirbu C., Oprea A., 2010 – Contribution to the Knowledge of the Alien Flora of Romania: Rudbeckia triloba L. and Senecio inaequidens DC. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 38(1): 33-36.
- Vladimirov V., Petrova A., 2009 – Senecio inaequidens (Asteraceae): a new alien species for the Bulgarian flora. Phytologia Balcanica, 15(3): 373-375.
Comportamento invasivo, strategie di invasività, Risk Assessment
- Bossdorf O., Lipowsky A., Prati D., 2008 – Selection of preadapted populations allowed Senecio inaequidens to invade Central Europe. Diversity and Distributions, Vol. 14, No. 4, 2008, pp. 676-685.
- Caño L., Escarré J., Sans F.X., 2007 – Factors affecting the invasion success of Senecio inaequidens and S. pterophorus in Mediterranean plant communities. Journal of Vegetation Science, 18 (2): 281-288.
- Garcia-Serrano H., Escarré J., Sans F.X., 2004 – Factors that limit the emergence and establishment of the related aliens Senecio inaequidens and S. pterophorus and the native S. malacitanus in Mediterranean climate. Canadian Journal of Botany 82: 1346-1355.
- Garcia-Serrano H., Sans F.X., Escarré J., Garnier E., 2005 – A comparative growth analysis between alien and native Senecio species with distinct distribution ranges. Ecoscience 12(1): 35-43.
- Lachmuth S., Durka W., Schurr F.M., 2010 – The making of a rapid plant invader: genetic diversity and differentiation in the native and invaded range of Senecio inaequidens. Mol Ecol., 19 (18): 3952-3967.
- Lachmuth S., Durka W., Schurr F.M., 2011 – Differentiation of reproductive and competitive ability in the invaded range of Senecio inaequidens: the role of genetic Allee effects, adaptive and nonadaptive evolution. New Phytologist, 192: 529-541.
- Lafuma, L., S. Maurice. 2007 – Increase in mate availability without loss of self-incompatibility in the invasive species Senecio inaequidens (Asteraceae). Oikos, 116: 201-208.
- López-García M.C., Maillet J., 2005 – Biological characteristics of an invasive south African species. Biological Invasions 7 (2): 181-194.
- Masante D., Barni E., Curtaz A., Bassignana M., Vidotto F., Tutino S., Siniscalco C., 2010. Past, present and potential distribution of the invasive Senecio inaequidens in the western Italian Alps (Aosta Valley). Proceedings of the 6th NEOBIOTA conference, Copenhagen (DK), 106.
- Monty A., 2009 – Sources de variation phénotypique des traits d’histoire de vie d’une espèce invasive, Senecio inaequidens DC. (Asteraceae). Thèse de doctorat. Gembloux, Faculté Universitaire des Sciences Agronomiques. 18 pp., 1 fig., 1 tableau, 7 articles.
- Monty A., Bizoux J.P., Escarré J., Mahy G., 2013 – Rapid plant invasion in distinct climates involves different sources of phenotypic variation. PloS one 8(1): 1-10. e55627.
- Monty A., Maurice S., Mahy G., 2010 – Phenotypic traits variation among native diploid, native tetraploid and invasive tetraploid Senecio inaequidens DC. (Asteraceae). Biotechnol. Agron. Soc. Environ., 14(4): 627-632.
- Monty A., Mahy G. 2010 – Evolution of dispersal traits along an invasion route in the wind-dispersed Senecio inaequidens (Asteraceae). Oikos 119: 1563–1570.
- Monty A., Mahy G., 2009 – Clinal differentiation during invasion: Senecio inaequidens (Asteraceae) along altitudinal gradients in Europe. Oecologia, 159: 305-315.
- Monty A., Stainier C., Lebeau F., Pieret N., Mahy G., 2008 – Seed rain pattern of the invasive weed Senecio inaequidens (Asteraceae). Belgian Journal of Botany. 141(1): 51-63.
- Pace L., Tammaro F., 2001 – The Main Invasive Alien Plants in the Protected Areas in Central Italy (Abruzzo). In: Visconti G. et al., Global change and protected areas. Advances in Global Change Research, 9: 495-504. Kluwer academic publ.
- Prati D., Bossdorf O., 2004 – A comparison of native and introduced populations of the South African Ragwort Senecio inaequidens DC. in the field. In: Breckle S.W., Schweizer B., Fangmeier A. (eds.) Results of worldwide ecological studies, pp. 353-359. Verlag Günter Heimbach, Stuttgart.
- Vacchiano G., Barni E., Lonati M., Masante D., Curtaz A., Tutino S., Siniscalco C., 2013 – Monitoring and modeling the invasion of the fast spreading alien Senecio inaequidens DC. in an alpine region. Plant Biosystems,147(4): 1139-1147.
- Verlinden M., De Boeck H.J., Nijs I., 2014 – Climate warming alters competition between two highly invasive alien plant species and dominant native competitors. Weed Research 54, 234–244.
- Verlinden M., Van Kerkhove A., Nijs I., 2013 – Effects of experimental climate warming and associated soil drought on the competition between three highly invasive West European alien plant species and native counterparts. Plant Ecol., 214: 243–254.
Impatti, intossicazione animali e uomo
- Caño L., Escarré J., Vrieling K., Sans F.X., 2009 – Palatability to a generalist herbivore, defence and growth of invasive and native Senecio species: testing the evolution of increased competitive ability hypothesis. Oecologia, 159: 95-106.
- Castells E., Morante M., Goula M., Pérez N., Dantart J., Escolà A., 2013 – Herbivores on native and exotic Senecio plants: is host switching related to plant novelty and insect diet breadth under field conditions? Insect Conservation and Diversity. doi: 10.1111/icad.12064.
- Cheeke P.R., 1984. Comparative toxicity and metabolism of pyrrolizidine alkaloids in ruminant and non ruminant herbivores. Canadian Journal of Animal Science, 64 (5): 201-202.
- Jacquemart A.L., Vanparys V., Meerts P., 2013 – Generalist versus Specialist Herbivores on the Invasive Senecio inaequidens and a Native Related Species: What Makes the Difference? American Journal of Plant Sciences, 4: 386-394.
- Reinhardt F., Herle M., Bastiansen F., Streit B., 2003 – Economic Impact of the Spread of Alien Species in Germany. Research Report UFOPLAN-Ref. No. 201 86 211. Federal Environmental Agency. Berlin, Germany.
- Schreber C., Crawley M.J., Porembski S., 2003 – The effects of herbivory and competition on Senecio inaequidens DC. (Asteraceae), an invasive alien plant. Diversity and Distributions, 9: 415-423.
- Passemard B., 2005. Intoxications du cheval par les séneçons. Thèse pour obtenir le grade de Docteur Vétérinaire. Ecole Nationale Vétérinaire de Toulouse. 94 p.
- Passemard B., Priymenko N., 2007 – L’intoxication des chevaux par les séneçons, une réalité en France. Revue Médicine Véterinaire: 158 (8-9): 425-430:
- Sarcey G, Gault G & Lorgue G (1992) Les intoxications par les senecons chez les equides [Senecio intoxications in horses]. Point Veterinaire. 23 (141): 965-970.
- Stegelmeier B.L., 2011 – Pyrrolizidine Alkaloid–Containing Toxic Plants (Senecio, Crotalaria, Cynoglossum, Amsinckia, Heliotropium, and Echium spp.). Veterinary Clinics of North America: Food Animal Practice, 27 (2): 419-428.
- Wiedenfeld H., 2011b – Toxicity of Pyrrolizidine Alkaloids – a Serious Health Problem. Müsbed, 1(2): 79-87.
Tossicità in miele e polline
- Bifulco E., 2010 – Studio degli aspetti nutrizionali e tossicologici di mieli uniflorali. Tesi di dottorato in Tossicologia degli Alimenti e dell’Ambiente, Ciclo XXIII, a.a. 2009-2010, Università degli Studi di Cagliari.
- Boppré M., Colegate S.M., Edgar J.A., 2005 – Pyrrolizidine alkaloids of Echium vulgare honey found in pure pollen. Agric. Food Chem., 53(3): 594-600.
- Di Marco G., Canuti L., Impei S., Leonardi D., Canini, A., 2012 – Nutraceutical properties of honey and pollen produced in a natural park. Agricultural Sciences, 3(2): 187-200.
- Dübecke A., Beckh G., Lüllmann C., 2011 – Pyrrolizidine alkaloids in honey and bee pollen. Food additives and contaminants, 28(3): 348-358.
- Dübecke A., Beuerle T., Ronczka S., Speer K., Wessel P., Beckh G., Lüllmann C., 2012 – Collection of Pyrrolizidine Alkaloid Plants & Pollen relevant for Honey Production. Research project AiF 16223 BG. German Ministry of Economics and Technology and FEI (Forschungskreis der Ernährungsindustrie).
- Kast C., Dübecke A., Kilchenmann V., Bieri K., Böhlen M., Zoller O., Beckh G., Lüllmann C., 2014 – Analysis of Swiss honeys for pyrrolizidine alkaloids. Journal of Apicultural Research. 53(1): 75-83.
Aspetti tossicologici: alcaloidi, inquinamento alimentare, normative
- Alexander J., Benford D., Boobis A., Eskola M., Fink-Gremmels J., Fürst P., Heppner C., Schlatter J., van Leeuwen R.; Special Issue: Risk assessment of contaminants in food and feed. EFSA Journal, 2012 – 10(10): s1004, 12 pp. doi:10.2903/j.efsa.2012.s1004.
- Bicchi C., Rubiolo P., Frattini C., 1991 – Off-line supercritical fluid extraction and capillary gas chromatography of pyrrolizidine alkaloids in Senecio species. Journal of Natural Products, 54 (4): 941-945.
- Carvalho S., Macel M., Schlerf M., Moghaddam F.E., Mulder P.P.J., Skidmore A.K., van der Putten W.H., 2013 – Changes in plant defense chemistry (pyrrolizidine alkaloids) revealed through high-resolution spectroscopy. Journal of Photogrammetry and Remote Sensing, 80: 51-60.
- Crews C., Berthiller F., Krska R., 2010 – Update on analytical methods for toxic pyrrolizidine alkaloids. Anal. Bioanal. Chem., 396: 327-338.
- Dimande A.F.P., 2007 – The toxicity of Senecio inaequidens DC. Thesis, Magister Scientiae, Department of Paraclinical Sciences, Faculty of Veterinary Science, University of Pretoria.
- Dimande A.F.P., Botha C.J., Prozesky L., Bekker L., Rösemann G.M., Labuschagne L., Retief E., 2007 – The toxicity of Senecio inaequidens DC. Journal of the South African Veterinary Association, 78(3): 121-129. EFSA (European Food Safety Authority), 2007 – Opinion of the Scientific Panel on Contaminants in the Food Chain on a request from the European Commission related to Pyrrolizidine Alkaloids as undesirable substances in Animal Feeds. EFSA Journal, 447: 1-51.
- EFSA (European Food Safety Authority), Panel on Contaminants in the Food Chain (CONTAM), 2011 – Scientific Opinion on Pyrrolizidine alkaloids in food and feed. EFSA Journal, 9(11): 2406, 134 pp. doi:10.2903/j.efsa.
- Fu P.P., Xia Q., Lin G., Chou M.W., 2004 – Pyrrolizidine Alkaloids-Genotoxicity, Metabolism Enzymes, Metabolic Activation, and Mechanisms. Drug Metabolism Reviews, 36 (1): 1-55.
- Kast C., Mühlemann M., Dübecke, A., Beckh, G., Lüllmann, C., 2010 – Alcaloidi pirrolizidinici in natura e importanza per gli alimenti. L’Ape, 9-10: 8-11.
- Martinello M., Cristofoli C., Gallina A., Mutinelli F., 2014 – Easy and rapid method for the quantitative determination of pyrrolizidine alkaloids in honey by ultra performance liquid chromatography-mass spectrometry: An evaluation in commercial honey. Food Control 37, 146-152.
- Rösemann G.M., Botha C.J., Eloff J.N., 2014 – Distinguishing between toxic and non-toxic pyrrolizidine alkaloids and quantification by liquid chromatography–mass spectrometry. Phytochemistry Letters 8: 126-131.
- Stegelmeier B.L., Edgar J.A., Colegate S.M., Gardner D.R., Schoch T.K., Coulombe R.A., Molyneux R.J., 1999. Pyrrolizidine alkaloids plants, metabolism and toxicity. Journal of Natural Toxins 8 (1): 95-116.
- Stewart M.J., Steenkamp V., 2001 – Pyrrolizidine Poisoning: A Neglected Area in Human Toxicology. Therapeutic Drug Monitoring, 23: 698-708.
- Wiedenfeld H., 2011 – Toxicity of Pyrrolizidine Alkaloids – a Serious Health Problem. Müsbed, 1(2): 79-87.
- Wiedenfeld H., 2011. Plants containing pyrrolizidine alkaloids: toxicity and problems. Food Additives & Contaminants, 28, 3, p. 282-292.
Sitografia
- DAISIE http://www.europe-aliens.org/speciesFactsheet.do?speciesId=23664#
- EPPO http://www.eppo.int/INVASIVE_PLANTS/ias_lists.htm
- ISSG http://www.issg.org/database/species/ecology.asp?si=1458&lang=EN
- SISSI http://sissi.divulgando.eu/specie/completo/4
- CABI http://www.cabi.org/isc/datasheet/49557#tab1-nav
- http://www.infoflora.ch/assets/content/documents/neofite/inva_sene_ina_i.pdf
- http://www.infoflora.ch/it/flora/8530-senecio-inaequidens.html#map
- http://www.itmonline.org/arts/pas.htm
- http://www.henriettes-herb.com/PAs/PAs-toxicity.html
- http://www.bfr.bund.de/en/frequently_asked_questions_on_pyrrolizidine_alkaloids_in_food-187360.html
- http://www.efsa.europa.eu/it/press/news/111108a.htm?wtrl=01
- www.equinescienceupdate.co.uk/ ragwort1.htm
Bibliografia (ordine alfabetico)
- Alexander J., Benford D., Boobis A., Eskola M., Fink-Gremmels J., Fürst P., Heppner C., Schlatter J., van Leeuwen R., 2012 – Special Issue: Risk assessment of contaminants in food and feed. EFSA Journal, 10(10): s1004, 12 pp. doi:10.2903/j.efsa.2012.s1004.
- Allegrezza, M., Biondi, E., Formica, E. & Ballelli, S., 1997 – La vegetazione dei settori rupestri calcarei dell’Italia centrale. Fitosociologia, 32: 91-120.
- Anzalone B., 1976 – Il Senecio inaequidens DC. in Italia. Giornale Botanico Italiano, 110: 437-438.
- Bartolucci F., Conti F., Tinti D., Scassellati E., Di Santo D., 2007 – La flora del Parco Nazionale del Gran Sasso e Monti della Laga. Biogeographia, vol. XXVIII: 39-44.
- Bicchi C., Rubiolo P., Frattini C., 1991 – Off-line supercritical fluid extraction and capillary gas chromatography of pyrrolizidine alkaloids in Senecio species. Journal of Natural Products, 54 (4): 941-945.
- Bifulco E., 2010 – Studio degli aspetti nutrizionali e tossicologici di mieli uniflorali. Tesi di dottorato in Tossicologia degli Alimenti e dell’Ambiente, Ciclo XXIII, a.a. 2009-2010, Università degli Studi di Cagliari.
- Biondi E., Allegrezza M., Taffetani F., Ballelli S., Zuccarello V., 2002 – Excursion to the National Park of Gran Sasso and Monti della Laga. Fitosociologia, 39 (1): 43-90.
- Biondi E., Allegrezza M., Zuccarello V., 2005 – Syntaxonomic revision of the Apennine grasslands belonging to Brometalia erecti, and an analysis of their relationships with the xerophilous vegetation of Rosmarinetea officinalis (Italy). Phytocoenologia, 35(1): 129-163.
- Biondi E., Blasi C. (Eds.), 2009 – Manuale Italiano di interpretazione degli habitat della Direttiva 92/43/CEE. http://vnr.unipg.it/habitat/.
- Biondi, E., Ballelli, S., Allegrezza, M. & Zuccarello, V., 1995 – La vegetazione dell’ordine Brometalia erecti Br.-Bl. 1936 nell’Appennino (Italia). Fitosociologia 30: 3-45.
- Boppré M., Colegate S.M., Edgar J.A., 2005 – Pyrrolizidine alkaloids of Echium vulgare honey found in pure pollen. Agric. Food Chem., 53(3): 594-600.
- Bornkamm R., 2002 – On the phytosociological affiliations of an invasive species Senecio inaequidens in Berlin. Preslia, 74(4): 395-407.
- Bossdorf O., Lipowsky A., Prati D., 2008 – Selection of preadapted populations allowed Senecio inaequidens to invade Central Europe. Diversity and Distributions, 14(4): 676-685.
- Bouvet D., Selvaggi A., Siniscalco C., Soldano A., 2013 – Senecio inaequidens DC. In: Bouvet D. (ed.), Piante esotiche invasive in Piemonte. Riconoscimento, distribuzione, impatti. Museo Regionale di Scienze Naturali, Torino, 352 pp.
- Braun-Blanquet J., 1932 – Plant sociology. Mac Graw Hill Book Comp., New York.
- Brunel S., Branquart E., Fried G., Van Valkenburg J., Brundu G., Starfinger U., Buholzer S., Uludag A., Joseffson M., Baker R., 2010 – The EPPO prioritization process for invasive alien plants. EPPO Bulletin, 40: 407–422.
- Bundesinstitut für Risikobewertung. (2001). Analytik und Toxizität von Pyrrolizidinalkaloiden sowie eine Eineshätzung des gesundheitlichen Risikos durch deren Vorkommen in Honig. Opinion No 038/2011. Berlin, Germany: Bundesinstitut für Risikobewertung, Federal Institute of Risk Assessment. Available from http:// www.bfr.bund.de/cm/343/analytik-und-toxizitaet-von-pyrrolizidinalkaloiden.pdf.
- Caño L., Escarré J., Sans F.X., 2007 – Factors affecting the invasion success of Senecio inaequidens and S. pterophorus in Mediterranean plant communities. Journal of Vegetation Science, 18(2): 281-288.
- Caño L., Escarré J., Vrieling K., Sans F.X., 2009 – Palatability to a generalist herbivore, defence and growth of invasive and native Senecio species: testing the evolution of increased competitive ability hypothesis. Oecologia, 159: 95-106.
- Carrara Pantano A., Tosco U., 1959 – Una nuova avventizia per la flora italiana: Senecio reclinatus L. f. di origine sud-africana, nella campagna veronese. Mem. Mus. Civ. St. Nat. Verona, Verona, 7: 151-157.
- Carvalho S., Macel M., Schlerf M., Moghaddam F.E., Mulder P.P.J., Skidmore A.K., van der Putten W.H., 2013 – Changes in plant defense chemistry (pyrrolizidine alkaloids) revealed through high-resolution spectroscopy. Journal of Photogrammetry and Remote Sensing, 80: 51-60.
- Castells E., Morante M., Goula M., Pérez N., Dantart J., Escolà A., 2013 – Herbivores on native and exotic Senecio plants: is host switching related to plant novelty and insect diet breadth under field conditions? Insect Conservation and Diversity. doi: 10.1111/icad.12064.
- Cheeke P.R., 1984 – Comparative toxicity and metabolism of pyrrolizidine alkaloids in ruminant and non ruminant herbivores. Canadian Journal of Animal Science, 64(5): 201-202.
- Chichiricco G., Frizzi G., Tammaro R., 1979 – Numeri cromosomici per la Flora Italiana. Informatore Botanico Italiano, 11: 3-35.
- Chytrý M., Otýpková Z., 2003 – Plot sizes used for phytosociological sampling of European vegetation. Journal of Vegetation Science, 14: 563-570.
- Conti F., 1997 – Minuartia glomerata subsp. trichocalycina comb. & stat. nov. (Caryophyllaceae), a Central Apennine endemic. Willdenowia, 27: 73-79.
- Conti F., 1998 – An annotated checklist of the flora of the Abruzzo. Bocconea, 10: 1-276.
- Conti F., Bartolucci F., 2009 – Prime indagini relative al gruppo Galium lucidum (Rubiaceae) in Italia. In: Società Botanica Italiana, Gruppo per la floristica. Gruppi critici della Flora d’Italia. Comunicazioni. Università “La Sapienza” Roma, 30-31 ottobre 2009.
- Conti F., Bartolucci F., Catonica C., D’orazio G., Londrillo I., Manzi A., Tinti D., 2006 – Aggiunte alla flora d’Abruzzo. II° contributo. Inform. Bot. Ital., 38(1): 113-116.
- Conti F., Bartolucci F., Manzi A., Miglio M. & Tinti D., 2008 – Aggiunte alla Flora d’Abruzzo. III contributo. Ann. Mus. civ. Rovereto, Sez. Arch., St., Sc. Nat.,23 (2007): 127-140.
- Conti F., Manzi A., Tinti D., 2002 – Aggiunte alla flora d’Abruzzo. I° contributo. Inform. Bot. Ital., 34(1): 55-61.
- Crews C., Berthiller F., Krska R., 2010 – Update on analytical methods for toxic pyrrolizidine alkaloids. Anal. Bioanal. Chem., 396: 327-338.
- Curtaz A., Talichet M., Barni E., Bassignana M., Masante D., Pauthenet Y., Siniscalco C., 2011 – Specie esotiche invasive e dannose nei prati di montagna. Caratteristiche, diffusione e metodi di lotta. Institut Agricole Régional, Aosta.
- Di Marco G., Canuti L., Impei S., Leonardi D., Canini, A., 2012 – Nutraceutical properties of honey and pollen produced in a natural park. Agricultural Sciences, 3(2): 187-200.
- Di Pietro R., Copiz R. & Catonica C., 2007 – Sulla presenza di Vaccinium gaultherioides Bigelow in Italia centrale. Biogeographia 28: 275–285.
- Dimande A.F.P., 2007 – The toxicity of Senecio inaequidens DC. Thesis, Magister Scientiae, Department of Paraclinical Sciences, Faculty of Veterinary Science, University of Pretoria.
- Dimande A.F.P., Botha C.J., Prozesky L., Bekker L., Rösemann G.M., Labuschagne L., Retief E., 2007 – The toxicity of Senecio inaequidens DC. Journal of the South African Veterinary Association, 78(3): 121-129.
- Dübecke A., Beckh G., Lüllmann C., 2011 – Pyrrolizidine alkaloids in honey and bee pollen. Food additives and contaminants, 28(3): 348-358.
- Dübecke A., Beuerle T., Ronczka S., Speer K., Wessel P., Beckh G., Lüllmann C., 2012 – Collection of Pyrrolizidine Alkaloid Plants & Pollen relevant for Honey Production. Research project AiF 16223 BG. German Ministry of Economics and Technology and FEI (Forschungskreis der Ernährungsindustrie).
- EC (European Commission), 2007 – Interpretation Manual of European Union Habitats, vers. EUR 27.
- EFSA (European Food Safety Authority), 2007 – Opinion of the Scientific Panel on Contaminants in the Food Chain on a request from the European Commission related to Pyrrolizidine Alkaloids as undesirable substances in Animal Feeds. EFSA Journal, 447: 1-51.
- EFSA (European Food Safety Authority), Panel on Contaminants in the Food Chain (CONTAM), 2011 – Scientific Opinion on Pyrrolizidine alkaloids in food and feed. EFSA Journal, 9(11): 2406, 134 pp. doi:10.2903/j.efsa.
- EPPO, 2004 – Reporting Service, No. 08. 2004/119. Invasive plants: addition of Senecio inaequidens and two Ludwigia species to the EPPO Alert List. Paris.
- EPPO, 2006 – EPPO data sheet on Invasive Plants. Senecio inaequidens. Web version 2006-02-01 – doc 05-11836.
- EPPO, 2012 – EPPO prioritization process for invasive alien plants. EPPO Bulletin, 42: 463–474. doi: 10.1111/epp.2592.
- Ernst W.H.O., 1998 – Invasion, dispersal and ecology of the South African neophyte Senecio inaequidens in the Netherlands: from wool alien to railway and road alien. Acta Botanica Neerlandica, 47(1): 131-151.
- Fiori A., 1923-1929 – Nuova Flora Analitica d’Italia. Tip. Ricci, Firenze.
- Fu P.P., Xia Q., Lin G., Chou M.W., 2004 – Pyrrolizidine Alkaloids-Genotoxicity, Metabolism Enzymes, Metabolic Activation, and Mechanisms. Drug Metabolism Reviews, 36 (1): 1-55.
- Gallina A., 2014 – Miele ed alcaloidi pirrolizidinici: presenza, diffusione e rischio. Comunicazione orale al Seminario del Parco Nazionale Gran Sasso e Monti della Laga, San Colombo di Barisciano (AQ); 11 novembre 2014.
- Garcia-Serrano H., Escarré J., Sans F.X., 2004 – Factors that limit the emergence and establishment of the related aliens Senecio inaequidens and S. pterophorus and the native S. malacitanus in Mediterranean climate. Canadian Journal of Botany 82: 1346-1355.
- Garcia-Serrano H., Sans F.X., Escarré J., Garnier E., 2005 – A comparative growth analysis between alien and native Senecio species with distinct distribution ranges. Ecoscience 12(1): 35-43.
- Grellier S., Ward D., Janeau J.L., Podwojewski P., Lorentz S., Abbadie L., Valentin C., Sébastien Barot S., 2013 – Positive versus negative environmental impacts of tree encroachment in South Africa. Acta Oecologica, 53: 1-10.
- Harland S.C., 1954 – The genus Senecio as a subject for cytogenetic investigation. Proceedings of the Botanical Society of the British Isles, 1, 256.
- Heger T, Böhmer H.J., 2005 – The invasion of Central Europe by Senecio inaequidens DC. – A complex biogeographical problem. Erdkunde, 59, 34-49.
- Heger T., Böhmer H.J., 2006 – NOBANIS – Invasive Alien Species Fact Sheet – Senecio inaequidens. – From: Online Database of the European Network on Invasive Alien Species – NOBANIS www.nobanis.org. Date of access 20/08/2014.
- Hijmans R.J., Cameron S.E., Parra J.L., Jones P.G., A. Jarvis, 2005 – Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25: 1965-1978.
- Hilliard O.M., Burtt B.L., 1975 – Notes on some plants of Southern Africa chiefly from Natal: IV. Notes from the Royal Botanic Garden Edinburgh, 34: 73-100.
- Jacquemart A.L., Vanparys V., Meerts P., 2013 – Generalist versus Specialist Herbivores on the Invasive Senecio inaequidens and a Native Related Species: What Makes the Difference? American Journal of Plant Sciences, 4: 386-394.
- Kast C., Dübecke A., Kilchenmann V., Bieri K., Böhlen M., Zoller O., Beckh G., Lüllmann C., 2014 – Analysis of Swiss honeys for pyrrolizidine alkaloids. Journal of Apicultural Research. 53(1): 75-83.
- Kast C., Mühlemann M., Dübecke, A., Beckh, G., Lüllmann, C., 2010 – Alcaloidi pirrolizidinici in natura e importanza per gli alimenti. L’Ape, 9-10: 8-11.
- Lachmuth S., Durka W., Schurr F.M., 2010 – The making of a rapid plant invader: genetic diversity and differentiation in the native and invaded range of Senecio inaequidens. Mol Ecol., 19 (18): 3952-3967.
- Lachmuth S., Durka W., Schurr F.M., 2011 – Differentiation of reproductive and competitive ability in the invaded range of Senecio inaequidens: the role of genetic Allee effects, adaptive and nonadaptive evolution. New Phytologist, 192: 529-541.
- Lafuma L., Balkwill K., Imbert E., Verlaque R., Maurice,S., 2003 – Ploidy level and origin of the European invasive weed Senecio inaequidens (Asteraceae). Plant Systematics & Evolution. 243(1-2): 59-72.
- Lafuma, L., S. Maurice. 2007 – Increase in mate availability without loss of self-incompatibility in the invasive species Senecio inaequidens (Asteraceae). Oikos, 116: 201-208.
- Le Bagousse-Pinguet Y., Forey E., Touzard B., Michalet R., 2013 – Disentangling the effects ofwater and nutrients for studying the outcome of plant interactions in sand dune ecosystems. Journal of Vegetation Science 24: 375-383.
- López-García M.C., Maillet J., 2005 – Biological characteristics of an invasive south African species. Biological Invasions 7 (2): 181-194.
- Macel M., Vrieling K., 2003 – Pyrrolizidine alkaloids as oviposition stimulants for the Cinnabar Moth, Tyria jacobaeae. Journal of Chemical Ecology, 29, (6): 1435-1446.
- Mattocks, A. R., 1986 – Chemistry and toxicology of pyrrolizidine alkaloids. London: Academic Press.
- Martinello M., Cristofoli C., Gallina A., Mutinelli F., 2014 – Easy and rapid method for the quantitative determination of pyrrolizidine alkaloids in honey by ultra performance liquid chromatography-mass spectrometry: An evaluation in commercial honey. Food Control, 37: 146-152.
- Masante D., Barni E., Curtaz A., Bassignana M., Vidotto F., Tutino S., Siniscalco C., 2010. Past, present and potential distribution of the invasive Senecio inaequidens in the western Italian Alps (Aosta Valley). Proceedings of the 6th NEOBIOTA conference, Copenhagen (DK), 106.
- Michez J.M., 1994 – Senecio inaequidens. From the wool to the vine. Phytoma, 468, 39-41.
- Monty A., 2009 – Sources de variation phénotypique des traits d’histoire de vie d’une espèce invasive, Senecio inaequidens DC. (Asteraceae). Thèse de doctorat. Gembloux, Faculté Universitaire des Sciences Agronomiques. 18 pp., 1 fig., 1 tableau, 7 articles.
- Monty A., Bizoux J.P., Escarré J., Mahy G., 2013 – Rapid plant invasion in distinct climates involves different sources of phenotypic variation. PloS one 8(1): 1-10. e55627.
- Monty A., Mahy G., 2009 – Clinal differentiation during invasion: Senecio inaequidens (Asteraceae) along altitudinal gradients in Europe. Oecologia, 159: 305-315.
- Monty A., Mahy G., 2010 – Evolution of dispersal traits along an invasion route in the wind-dispersed Senecio inaequidens (Asteraceae). Oikos 119: 1563–1570.
- Monty A., Maurice S., Mahy G., 2010 – Phenotypic traits variation among native diploid, native tetraploid and invasive tetraploid Senecio inaequidens DC. (Asteraceae). Biotechnol. Agron. Soc. Environ., 14(4): 627-632.
- Monty A., Stainier C., Lebeau F., Pieret N., Mahy G., 2008 – Seed rain pattern of the invasive weed Senecio inaequidens (Asteraceae). Belgian Journal of Botany. 141(1): 51-63.
- Muller S., Maillet J., 2004 – Senencio inaequidens DC. Le séneçon du Cap. In: Muller S. (coord.), Plantes invasives en France. Muséum national d’Histoire naturelle, Paris, 168 pp.
- Often A., 1997 – Senecio inaequidens DC. and Solidago rugosa Mill. found as ruderals in Oslo.- Blyttia, 55(3), 141-144.
- Pace L., Tammaro F., 2001 – The Main Invasive Alien Plants in the Protected Areas in Central Italy (Abruzzo). In: Visconti G. et al., Global change and protected areas. Advances in Global Change Research, 9: 495-504. Kluwer academic publ.
- Passemard B., 2005. Intoxications du cheval par les séneçons. Thèse pour obtenir le grade de Docteur Vétérinaire. Ecole Nationale Vétérinaire de Toulouse. 94 p.
- Passemard B., Priymenko N., 2007 – L’intoxication des chevaux par les séneçons, une réalité en France. Revue Médicine Véterinaire: 158 (8-9): 425-430.
- Pignatti S., 1982 – Flora d’Italia. Edagricole, Bologna.
- Pirone G., 2000 – La vegetazione ripariale nei versanti nord-orientali del Gran Sasso e dei Monti della Laga (Abruzzo, Italia). Fitosociologia, 37(2): 65-86.
- Pirone G., Frattaroli A.R., Biondi E., Casavecchia S., Pesaresi S., 2010 – La vegetazione forestale del Parco Nazionale del Gran Sasso e Monti della Laga. L’Italia Forestale e Montana. 65(6): 699-735.
- Prati D., Bossdorf O., 2004 – A comparison of native and introduced populations of the South African Ragwort Senecio inaequidens DC. in the field. In: Breckle S.W., Schweizer B., Fangmeier A. (eds.) Results of worldwide ecological studies, pp. 353-359. Verlag Günter Heimbach, Stuttgart.
- Rameau J.C., Mansion D., Dumé G., 1989 – Flore Forestiere Francaise, Tome 1: Plaines et Collines. Institute pour le Developpment Forestier.
- Rameau J.C., Mansion D., Dumé G., 1993 – Flore Forestiere Francaise, Tome 2: Montagnes. Institute pour le Developpment Forestier.
- Rameau J.C., Mansion D., Dumé G., Gauberville C., 2008 – Flore Forestiere Francaise, Tome 3: Region Mediterranéenne. Institute pour le Developpment Forestier.
- Reinhardt F., Herle M., Bastiansen F., Streit B., 2003 – Economic Impact of the Spread of Alien Species in Germany. Research Report UFOPLAN-Ref. No. 201 86 211. Federal Environmental Agency. Berlin, Germany.
- Rösemann G.M., Botha C.J., Eloff J.N., 2014 – Distinguishing between toxic and non-toxic pyrrolizidine alkaloids and quantification by liquid chromatography–mass spectrometry. Phytochemistry Letters 8: 126-131.
- Sans F.X.; Garcia-Serrano H.; Afán I., 2004 – Life-history traits of alien and native senecio species in the Mediterranean region. Acta Oecologica, 26 (3): 167-178.
- Sarcey G., Gault G., Lorgue G., 1992 – Les intoxications par les senecons chez les equides. Point Veterinaire, 23(141): 965-970.
- Schmitz G., Werner D.J., 2001 – The importance of the alien plant Senecio inaequidens DC. (Asteraceae) for phytophagous insects. Zeitschrift für ökologie und Naturschutz, 9(3), 153-160.
- Schreber C., 2002 – The effects of herbivory and competition on Senecio Inaequidens DC. (Asteraceae), an invasive alien plant. Diploma Thesis, University of Rostock., 92 pp.
- Schreber C., Crawley M.J., Porembski S., 2003 – The effects of herbivory and competition on Senecio inaequidens DC. (Asteraceae), an invasive alien plant. Diversity and Distributions, 9: 415-423.
- Sirbu C., Oprea A., 2010 – Contribution to the Knowledge of the Alien Flora of Romania: Rudbeckia triloba L. and Senecio inaequidens DC. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 38(1): 33-36.
- Stegelmeier B.L., 2011 – Pyrrolizidine Alkaloid–Containing Toxic Plants (Senecio, Crotalaria, Cynoglossum, Amsinckia, Heliotropium, and Echium spp.). Veterinary Clinics of North America: Food Animal Practice, 27 (2): 419-428.
- Stegelmeier B.L., Edgar J.A., Colegate S.M., Gardner D.R., Schoch T.K., Coulombe R.A., Molyneux R.J., 1999. Pyrrolizidine alkaloids plants, metabolism and toxicity. Journal of Natural Toxins 8(1): 95-116.
- Stewart M.J., Steenkamp V., 2001 – Pyrrolizidine Poisoning: A Neglected Area in Human Toxicology. Therapeutic Drug Monitoring, 23: 698-708.
- Svenning J.-C., Gravel D., Holt R.D., Schurr F.M., Thuiller W., Münkemüller T., Schiffers K.H., Dullinger S., Edwards T.C., Hickler T., Higgins S.I., Nabel J.E.M.S., Pagel J., Normand S., 2014 – The influence of interspecific interactions on species range expansion rates. Ecography 37: 1-12. doi: 10.1111/j.1600-0587.2013.00574.x.
- Tammaro F., 1995 – Lineamenti floristici e vegetazionali del Gran Sasso meridionale. Documenti naturalistici per la conoscenza del Parco Nazionale del Gran Sasso-Laga. Boll. Mus. Civ. St. Nat. Verona, 19(1992): 1-256.
- Thuiller W., Georges D., Robin R., 2014 – biomod2: Ensemble platform for species distribution modeling. R package version 3.1-48. http://CRAN.R-project.org/package=biomod2.
- Tutin T.G. & al., 1964-1980 – Flora Europaea. Vol. 1-5. Cambridge University Press, London.
- USDA, 2005 – Weed Risk Assessment for S. inaequidens DC. and S. madagascariensis Poir (Asteraceae). United States Department of Agriculture, Animal and Plant Health Inspection Service.
- USDA-APHIS, 2013 – Weed Risk Assessment for Senecio angulatus L. f. (Asteraceae) – Cape-ivy. Version 1. United States Department of Agriculture, Animal and Plant Health Inspection Service.
- Vacchiano G., Barni E., Lonati M., Masante D., Curtaz A., Tutino S., Siniscalco C., 2013 – Monitoring and modeling the invasion of the fast spreading alien Senecio inaequidens DC. in an alpine region. Plant Biosystems, 147(4): 1139-1147.
- Vanparys V., Cawoy V., Mahaux O., Jacquemart A.-L., 2011 – Comparative study of the reproductive ecology of two co-occurring related plant species: the invasive Senecio inaequidens and the native Jacobaea vulgaris. Plant Ecology and Evolution, 144(1): 3-11.
- Vanparys V., Meerts P., Jacquemart A.-L., 2008 – Plant–pollinator interactions: comparison between an invasive and a native congeneric species. Acta Oecologica, 34: 361-369.
- Verlinden M., De Boeck H.J., Nijs I., 2014 – Climate warming alters competition between two highly invasive alien plant species and dominant native competitors. Weed Research, 54: 234-244.
- Verlinden M., Van Kerkhove A., Nijs I., 2013 – Effects of experimental climate warming and associated soil drought on the competition between three highly invasive West European alien plant species and native counterparts. Plant Ecol., 214: 243–254.
- Vladimirov V., Petrova A., 2009 – Senecio inaequidens (Asteraceae): a new alien species for the Bulgarian flora. Phytologia Balcanica, 15(3): 373-375.
- Wiedenfeld H., 2011a – Plants containing pyrrolizidine alkaloids: toxicity and problems. Food Additives & Contaminants, 28(3): 282-292.
- Wiedenfeld H., 2011b – Toxicity of Pyrrolizidine Alkaloids – a Serious Health Problem. Müsbed, 1(2): 79-87.
- Wittenberg R. (ed.), 2005 – An inventory of alien species and their threat to biodiversity and economy in Switzerland. CABI Bioscience Switzerland Centre report to the Swiss Agency for Environment, Forests and Landscape.